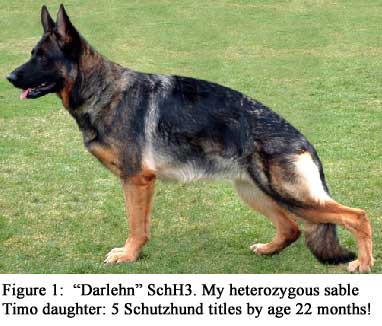
Most German Shepherd Dogs (the full name of the breed) are colored, and these are listed as: 1. sable (see Fig.1), 2. black-and-tan (let’s call this one “B&T” referring to a dog with a black saddle, Fig.2), 3. black-and-tan of the Rottweiler-type coloration (usually called “bi-color”), and 4. solid black–including black hair under the tail around the anus (Fig. 4). In addition, a 5th color is white, which is not given equal privileges by most registries around the world, and consequently finds itself most of the time in some isolation regarding show competitions. See Fig. 5
The white dog has been in the population since before the beginning of the German Shepherd Dog; that is, the color variety has existed since before recognition as a distinct breed. When the GSD began to be recorded (registered, identified by name and number) and an association formed for that purpose a decade before 1900, there were various “styles” or types used by sheep herders and other owners. There were heavy-bodied dogs with hanging ears similar to those of the Pyrenean flock guardian, and light-weight dogs with bodies and erect ears similar to Spitz types. There were what would later become known in English as “sables” but in German as “grey” (grau), this being the most populous for many decades. There were some dogs with black saddles and brown or tan legs (schwartz und gelb) in a wide range of just how much of each covered the body. There were fewer with so much black covering the torso and legs that the impression of color was the same as that seen in the Rottweiler and several other breeds already established or in process of selection. There were a few of the solid black dogs, some with a tiny presence of some brown hairs on feet or haunches, some with none at all.
And, there is also the white dog. The all-white GSD seems to have been in the gene pool as early as any other color variation, but since the founder of the breed club, Max von Stephanitz, had some erroneous ideas about inheritance, it was considered a sign of weakness, and soon was banned from the breeding registry and show rings of the Deutsche Schäferhund. However, the genes lay hidden in the mainstream and many of the dogs remained in the field or yard, doing what their colored “brothers and sisters” did. In scattered locations, where novelty was appreciated, they were even bred intentionally. The development of cadres of fanciers was found in the homeland and eventually in other countries.
The white German Shepherd Dog had its greatest acceptance and population in the United States, the country with the largest number of people and room for dogs that were not exclusively bred and kept for work. While a few of these did find jobs as blind-guide and other-service dogs, most were family pets. And despite the ban by the German breed club, the Schäferhund Verein (SV), white dogs for a few years continued popping up in litters produced by colored parents. When the Second World War began (and the population of GSDs in private hands fell precipitously), white GSDs in Europe were nearly extinct. However, the bank in America was well-stocked, and (except for a handful in England) was the only place where they could be found. It was this gene pool that eventually repopulated this color variety around the world.
Today, the white German Shepherd Dog is not “recognized” by most national kennel clubs around the world, because most are affiliated with or influenced by the true “parent club”, the SV, in Germany. Even in those countries in which the major “national kennel club” has maintained more independence from FCI, SV, etc. (such as the USA, the UK, Australia, and a very few others), they generally follow the German (SV) breed standard guidelines to some extent. These consider white coats to be a show (or even a registration) disqualification.
Today there are fairly large populations of white GSDs in a great number of countries, most notably the USA, but also good numbers in the UK and the rest of Europe, and fewer in other countries (though still more numerous than many breeds). Some have established the white dog as a separate breed, regardless of the same heritage. Others include it as an equal member in regard to interbreeding and exhibition. In all of these various communities, the genetics of coat pigment in this color variety are mostly misunderstood.
A few writers have tried to explain or describe the white GSD, some from the viewpoint of history and pedigrees and individuals. But only a couple of us, to my knowledge, have delved into the genetics. Dr. Malcolm Willis, a professional geneticist, is one; I am another. My late friend Malcolm had the advantage of much study in several species, while I have the advantage of having had contact with vastly greater numbers of examples and exceptions due to my living in the USA, and to a lesser extent from having judged dog shows in about 30 countries. In all, I was always on the alert for genetic evidence. While Malcolm also traveled abroad, he had most of his observation opportunities in that smaller-population island, Great Britain. He had not seen the numbers and variations that I have been exposed to, and for most of his career had been hampered somewhat by the UK’s quarantine policy.
There are two major reasons why the inheritance of the white GSD has long been a mystery, but I believe I have solved the bigger part of the puzzle. One factor is that the white dog is in the minority and thus its genetics have not been a primary serious subject of interest by what I call “practical geneticists”—breeders. The other is that breeders of varieties that are not universally accepted (sometimes considered “outcasts”) are less likely to be experienced cynologists. That is, they are less likely to include scientific geneticists, and simply are in the game for fun and their love of dogs. The students of genetics are more likely to be found among the mainstream of dog fanciers. Additionally, it takes time away from other pursuits. Had I been a breeder of white Shepherds all my years in dogs, I would have studied the genetics in greater depth long ago. Since I retired, I have spent more time in judging, researching, and writing.
It had long been thought by the vast majority of fanciers that the white GSD was always a recessive color variety. But it seems that to date, no one had clearly explained how or why. No one, to my knowledge, had published information (much less explained to the dog public) that sables (a dominant trait and thus requiring at least one parent to be sable) could result from a pairing of dogs neither of which had a recognizable sable color pattern. I felt it was time to propose a cogent and illustrated theory aimed toward such understanding.
I have judged the White Shepherd in many shows, including numerous times at the national specialty of the American club for the white GSD, as well as at United Kennel Club shows. I judged the pan-European White Shepherd International show that was held in Holland as well. When I was “on the road” in my occupation as technical advisor for a chemical company, I also had opportunity to visit kennels large and small that specialized in breeding the white dog.
As a pro handler in their part of the country, I was “in on the ground floor” during the birth and growth of the “Shiloh Shepherd,” an offshoot of the GSD whose fanciers specialized in large size, but also blended many sables and whites into their pool. I have judged their national specialty show numerous times, also, and have kept informed as to their peculiar genetics. Regardless of your interest in the Shiloh Shepherd, this movement has provided much grist for the genetics mill. In my functions as consultant and judge for such groups, I have had access to more information than has befallen most observers of the canine scene, and some useful data has come from the Shiloh breeding community.
This Shiloh population came about by a selection process in the larger GSD field several decades ago, and has been segregated long enough and thoroughly enough to be considered an independent breed. However, that does not materially change genetics. Some years ago the original club (ISSR) added white dogs to the gene pool of Shiloh Shepherds (which had already included a preponderance of sables). A couple of other registries also allow the breeding of white to colored dogs but have not changed the breed name. For the sake of convenience as well as clarity regarding genetics, I will refer to them all as GSDs.
In this discussion of the genetics of the white GSD, we will not include the color of the nose bulb or other “integument” because pigment in the nails, nose and pad leather, eyerims, lips, and anus is generally unrelated to coat pigment inheritance. Nor will the subject of “white markings” be covered here, as that is also a separate genetic pathway we can walk at another time.
Inherited characteristics are determined by strands of DNA that we call chromosomes, and the genes that are clumped along those strands. Some genes are found on certain chromosomes, other genes are on different ones. It seems that all or most of the genes determining a dog’s color phenotype are on one chromosome. At least, inheritance works in such a way that we can assume so without risk. Some genes determine leg length, others head shape, etc., but we are concerned with a few that dictate what general color pattern and hue our dogs are. It is convenient to give types of genes certain letters, sometimes arbitrary and sometimes standing for something.
Assume that a particular chromosome strand contains all the genes for coat color. Imagine, for purposes of illustration, that genes look like individual knots tied in this string. We give them letter-names designating what characteristics those knots call for. The places where those genes are, we call loci (plural for locus, or location). On the coat-color chromosome, we say that the gene for pattern (sable, saddle, bi-color B&T) is at the A locus. There can be a choice of what sub-type of gene is at that locus, so we have to distinguish between them. The most dominant allele (version or variety of that particular gene) is normally capitalized. Thus, we give the capital-letter “A” to the dominant pattern gene. In the GSD this is “sable,” meaning a relatively uniform all-over coloration distinguished by black hair tips so the dog looks like its light- or medium-shaded coat has been stroked with a soot-covered cloth or glove. Sables usually have much less black tipping on the legs and underparts than on the neck, back, and sides. Especially the heterozygous ones. To designate that we have the sable gene at the A locus, we use a superscript “y”, which is placed upward a little, like where an exponent or apostrophe would be. If the printer or computer this article is sent to can “read” my superscripts, it will look like this: Ay. That means that at the A locus, there is the configuration of the DNA that calls for sable.
Genetic dominance should not be equated with population dominance; i.e., a “stronger” characteristic may actually be seen less often, such as when the “weaker” characteristic has been selectively bred for by people. Nature follows other rules, which is why recessive traits are less-often seen in the wild. Which is the main reason why there are far more “sable” or “wolf-gray” wild canids seen in wolf packs and coyote families than solid blacks or solid whites.
The A-locus gene calling for the saddle-marked B&T that is the most commonly seen coat pattern in GSDs is given the designation as. The “s” could mean “saddle.” If you see at, you can remember this “t” as meaning “tan points”—a “bi-color” dog with Rottweiler type markings. We’ll leave the solid black pattern for later.
Now, you need to remember that nearly all cells have two genes of the same general type in each locus. The exception is in the sex cells, sperm and ovum, which each carry only one of each pair. Otherwise the number would double with every impregnation! If both members of the pair are the same, the animal is said to be homozygous for that trait; if the pair is made up of two slight variations, it is heterozygous. A homozygous sable has AyAy at this locus, meaning it cannot contribute to its offspring any pattern gene other than Ay. If the pup is to get something else for the other half of its genotype, it will have to come from the other parent. A dog that has Ayas is still going to be a sable in appearance, though perhaps not as strikingly… the recessive gene, from the other as (B&T) or at (bi-color) parent might have just enough influence to “show through” a little and give a hint of its genotype presence (not always, though). There are some heterozygous sables that have so much black tipping on the back that people wonder if it is a black-and-tan with a lot of “grizzling” or lighter undercoat (or the lower part of topcoat hairs) showing. Others have much less, and can be mistaken for a faded B&T. It sometimes helps to see the parents to get a clue.
A different coat-color chromosome locus is given the E designation, and you can consider that to stand for “extension” or “expression” of color in the hair. Usually a capital letter is used for the most dominant member of each gene family. The E locus either demands (EE), allows (Ee), or denies (ee) expression of color in the hair. In other words, these alleles affect the production or non-production of pigment granules in the medullae (centers) of the hair shafts. No matter what the dog has at the A locus, the rank of the E traffic cop at the corner is going to tell the A-locus genes whether or not they will be allowed onto the highway (become visible). So you can have a dominant coat-color pattern (sable, which means “black tips”) but an “ee” cop that doesn’t allow it any access, so what do you have? A dog with no color in the hairs. A white dog. A “white sable.”
This prohibition of coloration is not always 100 percent, because of the interactions of what we call “modifying genes” on these main genes. So, fairly frequently we see a white dog with “biscuit” (very light tan) coloration on ears, withers, and perhaps along the spine a ways. Don’t worry about modifier genes on distant loci unless you are breeding strictly for purity of color. I have even seen all-buff (very light tan) individuals.
A dog, whether sable, saddle, or bi-color (Ay, as, or at) that has a double dose of the e allele (at the E locus) calling for “no expression” of color in hair will be a white dog. If it is a color-suppressed sable (Ayee), it will be a dominant white, because the sable Ay pattern is dominant over other patterns. If both A-locus alleles are Ay, all of that homozygous dog’s offspring will be sables, whether colored or white. If white due to the ee presence on that other locus, such a dog might be a bright or “clean” white, if unidentified modifiers elsewhere have an effect. Remember, “sable” is really basically a pattern, not a color. Color is dictated by genes at loci other than A, and the A-locus genes just direct the color (if present) where to appear. It is quite possible that all white GSDs are actually sables with the ee allele. To my knowledge, nobody has attempted a genetic experiment to find out if this is the case, or if other-pattern GSDs can also be ee-white.
Put it another way: if the white dog you are looking at has no dominant sable Ay allele, it is either as or at, at that locus. There is no way you can be sure without a color pedigree and probably a test mating, or data from several different matings of the parents. Even then, it may be just a guess.
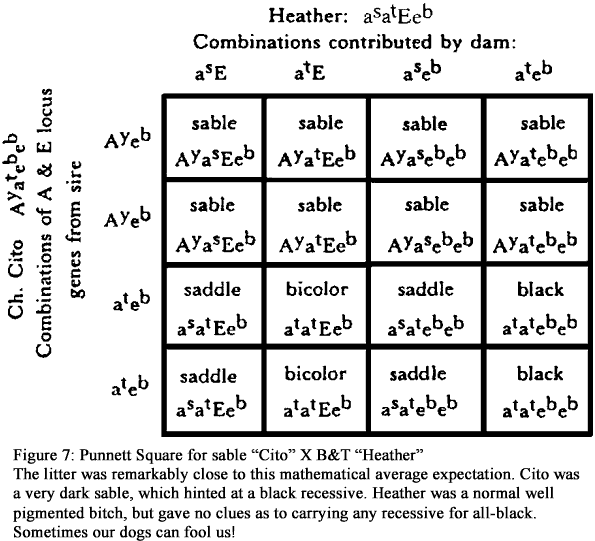
Now, some readers have undoubtedly wondered about the relation of solid black GSDs to the other patterns. It is almost certain that the determinant for solid black coat color is on a different location than for the “other” patterns. Or, more accurately, a result of cooperation between genes on different loci. This other allele that we believe can possibly exist at the E locus is given the designation eb. If this is indeed where it lies, then it is the most recessive of the E class. In double dose, it is responsible for the all-black GSD, but it seems that in order to express itself, it must have some “help” from the double presence of at on the A locus. This conclusion is reached because some litters have sables, saddle-B&Ts, bi-colors, and all-blacks in the same batch. The Punnett square for Heather (a B&T whose phenotype gave no clue to a black recessive) bred to Cito (a very dark sable, hinting of the black factor) shows all four of these color variations. The doubling up of the atatebeb genes is the most likely explanation. See Figures 4 and 7.
It was once thought that bi-color dogs produced more solid blacks than other color patterns did, but that theory does not hold water once you learn that Bi X Bi (at bred to another at) gives a lower percentage of solid blacks than you would expect if the bi-color GSD was that way because of a black recessive in every case. Instead of 25%, one study showed it to be about 3% solid blacks coming from two bi-color B&Ts. No, the solid black does not come entirely from the A locus; the most we can say is that it is possible that the double presence of at there is needed in order for the double eb at the E locus to work. Some sort of chemical cooperation between E locus and A locus, if the “right” alleles are there. It only appears that the parents each need to carry a single recessive at gene, and when they get together in the presence of eb from each, you get a black pup with the genotype atatebeb.
The “bicolor” GSD (with Rottie-type markings) has some variation in phenotype. Probably most have some tan markings on cheeks, bib, and eyebrow dots, and black on the back of the metatarsus (extending down from the hock joint). Perhaps those with almost completely black faces and/or black past the hocks have a recessive for solid black in the form of an atatEeb genotype. Sometimes, especially when the black color does not extend quite so far down the legs, it is hard to guess whether the individual is a bicolor with a little less expression, or a bicolor with that black recessive mentioned in the previous sentence, or a heavily-marked non-bicolor of asatebeb or asatEeb makeup. See Figures 3a and b.
A Shiloh litter was brought to my attention that had a white-looking sire, “Scout” and a saddle-marked B&T dam, “Elf”. However, the litter included some B&T whelps and some sable whelps, all with normal color as you might see in any litter produced by a heterozygous sable bred to a B&T. Now, the only way you can get sables is to have a sable parent (or two). So the breeder and friends were confused, and wrote to me for an explanation. I told them that, to a geneticist, the answer was “plain as the nose on your face” though not obvious to those unfamiliar with the science. The “white” dog (and it is indeed phenotypically white, as I can see only a narrow, faded light-cream band on its back and a little shade of cream-tan on the back of the ears) is genetically a sable whose black hair pigment has been inhibited from ever developing because of that “ee” traffic-cop pair of genes. See figure 6.
Another set of pictures of Shilohs (bottom of Figure 6) shows the “Gem/Quest” litter born to a light-colored sable and its B&T as (saddle) partner that appears to also have the at (bi-color) recessive. It’s only a guess, based on how much black covers the dog, that it probably has the heterozygous asat genotype rather than homozygous atat. And one pup had pretty typical bi-color markings such as the tan eyebrow dots. Some of the pups were typical (colored) sables, as would be expected due to the fact that sable is a dominant pattern and is usually expected when one parent is. However, remember that the distribution (percentages of each pattern) is an average, and it is possible that one litter can be all sables and the next litter from the same dogs can be all B&Ts. If the sable parent had been homozygous for that pattern, all of the litter would have been sable-patterned.
Colors of the offspring varied, as you see in the accompanying illustration. B&Ts ranged from no mask to partial mask to what appears to be a bicolor dog (the one with the tan eyebrow “dots”), and others include a short-coat faded probable-sable and a darker long-coat sable.
But one pup was phenotypically white. Why? Neither parent had a phenotype of that color (white). The answer probably lies in the fact that each parent has one copy of the “e” allele at its E locus. That is, both were Ee there, and when you put two of those combinations together, you can get some EE, some ee, and more Ee. The white dog was ee, meaning that it had genes for not allowing expression of pigment. It is probably a genetic sable, but without color being expressed, one can’t be positive… it could as well be a B&T or a bi-color (because each parent carried one of these recessives), but since the ee gene refuses to allow color to be shown in the hair shafts, we don’t know for sure until it is given a test breeding.
After I had submitted the first (truncated) draft of this article to the editor of the Shiloh breed club newsletter, she sent this question: “Since dual [their club’s term for either at bi-color or as saddle-marked dog] is a recessive gene, how can such a dual dog carry the white recessive? Or is the white recessive at a different locus?”
She almost hit the nail squarely on the head. Yes, a different locus is also involved. Suppose the dog’s markings or color is a product of manufacturing with parts made or steps taken at various factories along a certain highway. The tan-point (bi-color) gene, the saddle-marking gene, and the sable gene are all found at the A locus (like Toledo on Interstate-75) but the extension genes such as E and e are found at the E locus (like Cincinnati on I-75). Cincinnati has the ability to ship (or deny shipment) to Toledo the authorization to include black (dark color) in the final product (the hair coat). Each city (locus) is only relatively independent of each other, as the cities on an Interstate are.
The genes for pattern (such as bi-color markings or saddle-B&T markings or sable) are in Toledo, and are only affected by the E/e genes (for: “Black expression, yes or no?”) made in Cincinnati, to the extent that the truck terminal and assembly plant in Cincinnati (E locus) does not allow any dark-pigment authorization to go to Toledo if they decide that the dog’s description is to be “ee”. The terminal manager at E denies the terminal and assembly plant at Toledo any dark-pigment approval, so only a colorless (white) product leaves the Toledo warehouse/terminal. That’s all Toledo can put into the product, therefore. A GSD/Shiloh of any color-pattern must have approval from the manager at the Cincinnati E locus (in the form of EE or Ee) before it can make dark pigment appear in the hairs. No approval (or only for ee) means no color (a white dog). Toledo releases product with color only when Cincinnati allows.
When white GSDs are brought into a colored family line, we frequently see offspring that almost look like “black-and-white” dogs because of sharp contrast between the dark upper parts (such as the saddle and head markings) and the lower markings or underparts that in other dogs are more often some shade of tan or brown—legs, belly, etc. This may hint at yet another chromosome/gene locus that is given the C designation. That is, many white Shepherds may have alleles at the C locus that call for paling pigment. Think of C standing for “Concentration.” Genes at this locus affect only the hair color in the lower parts of the coat or, to put it another way, those areas other than where the saddle and other dorsal portions would be. Only the “tan parts.” The most dominant of these alleles is given the capital-C designation. A less-dominant (recessive) allele, called “the chinchilla gene” because of superficial resemblance to members of that rodent group, is given the designation cch. Full (or nearly-full) expression of color in the “tan parts” is seen in homozygous CC dogs, but phenotypically, you might or might not see any difference in shade when a heterozygous Ccch dog is presented. However, when cchcch is present in double dose, the contrast between dark and light parts is eye-catching.
The “chinchilla gene” in the colored GSD has been selected against (bred out) to a great extent, although there are population pockets resulting from an extremely small number of breeders who fancy this dark-light combination. Also, it persists among those who breed whites or both white and colored dogs, and that is probably because its presence is invisible when the dog is white anyway. Also, most “nearly black-and-white” dogs are found to have whites in the near pedigree. A “fading gene” would not be noticed in a population of dogs without hair pigment, so this cch allele can be considered immaterial in white-dog populations.
In conclusion, we can say that the occurrence of all-white in GSDs (and many other breeds) comes from the double presence on the E-locus of the allele called “e”— that is, two copies of the gene calling for “no color expression.” It can be a genotypic sable or (perhaps) a genotypic other-pattern dog, but in any case, it is missing a dominant “extension” (E) allele. Breed a white dog to a B&T and, if sable pups are born, the former is a white sable.
Incidentally, but not that you are likely to come across this (except in the pages of my GSD book), there is also the color oddity of the all-over buff-color (tan) dog with no markings and almost no shadings. I have seen this when a white dog was mated to a solid black dog. It would give credence to the theory that the coding for solid black pattern might not be on the E locus alone, as had once been thought, and that the all-tan dogs might have been a result of ee (no pigment expression) interacting with a relatively “strong” gene (or genes) calling for all-over dark pigment expression, inherited from the solid blacks. There is also the apparently all-“buff” or tan-colored dog with self-color nose. This is a result of something else; namely, an otherwise-black dog with the double recessive “bb” allele for “liver.” Unless you look at the nose bulb, eyerims, anus, lips, etc., you might miss the fact that this is a liver dog, rather than a buff resulting from a white-X-black cross. There are still many questions to be answered, and we need accurate color-pedigree records plus someone to compile and analyze them.